


科学传播

大冰川中的小精灵—青藏高原冰川微生物
发布时间:
2023-03-06

巍峨雄伟的青藏高原上分布着南北极以外地球上面积最大的冰川,蜿蜒连绵,构成雪白纯洁晶莹透剔的冰雪世界景观。青藏高原冰川是亚洲水塔的固体水库,储存着南北极外最大量的固态淡水;青藏高原冰川也是过去地质和历史时期微生物的天然存储库,封存了季风环流搬运、西风环流搬运和局地生长的不同种类微生物。青藏高原是西风带与亚洲季风环流系统交汇区,所以季风环流搬运的微生物来自海洋及沿途不同生态系统的微生物,西风环流搬运的微生物则源于欧洲及中亚等地区。
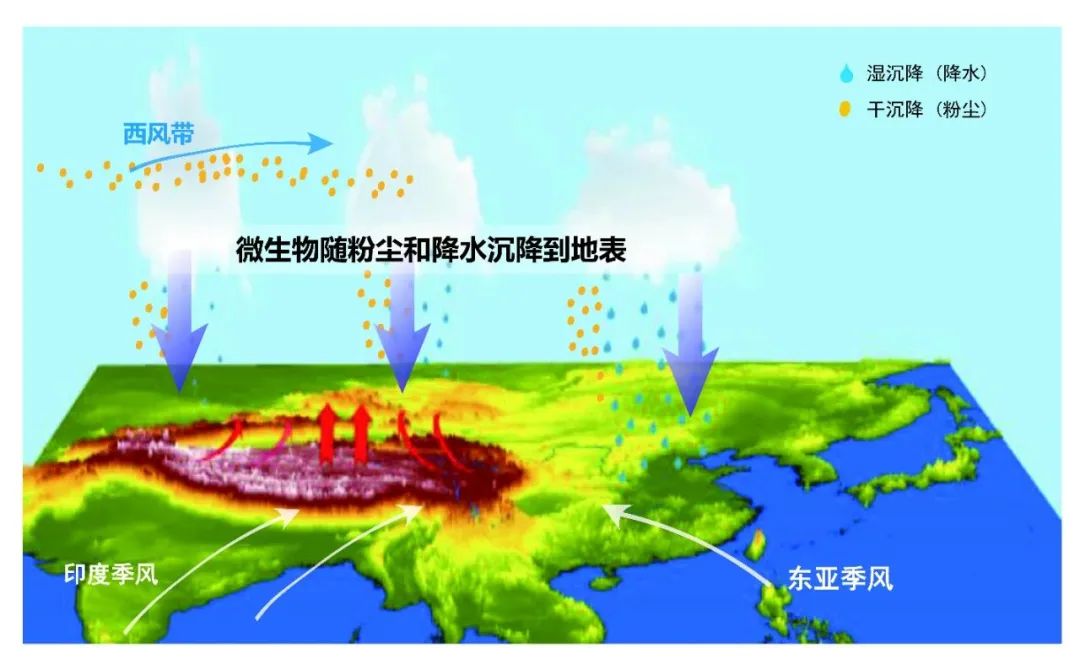 图3. 西风季风带来不同生态系统的微生物沉降到青藏高原,图源:第二次科考队。
图3. 西风季风带来不同生态系统的微生物沉降到青藏高原,图源:第二次科考队。

图5.珠穆朗玛峰东绒布冰川:是建立青藏高原冰川微生物基因组和基因数据集的典型冰川,图源:第二次科考队。

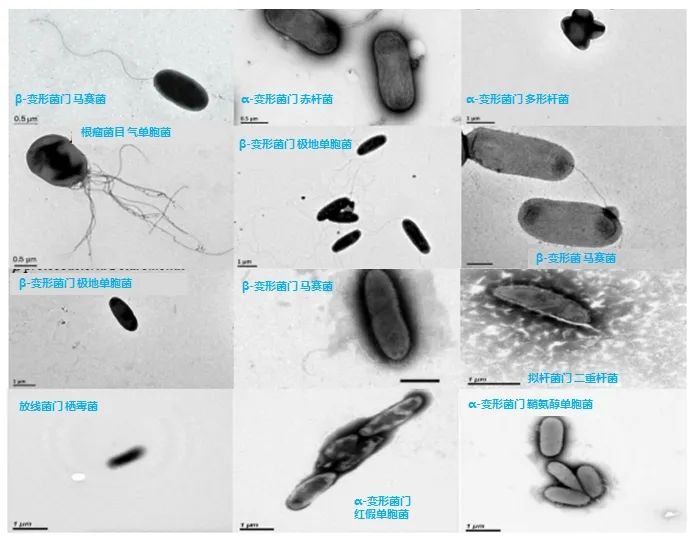
图7. 从青藏高原冰川中分离培养的细菌新种,图源:第二次科考队。
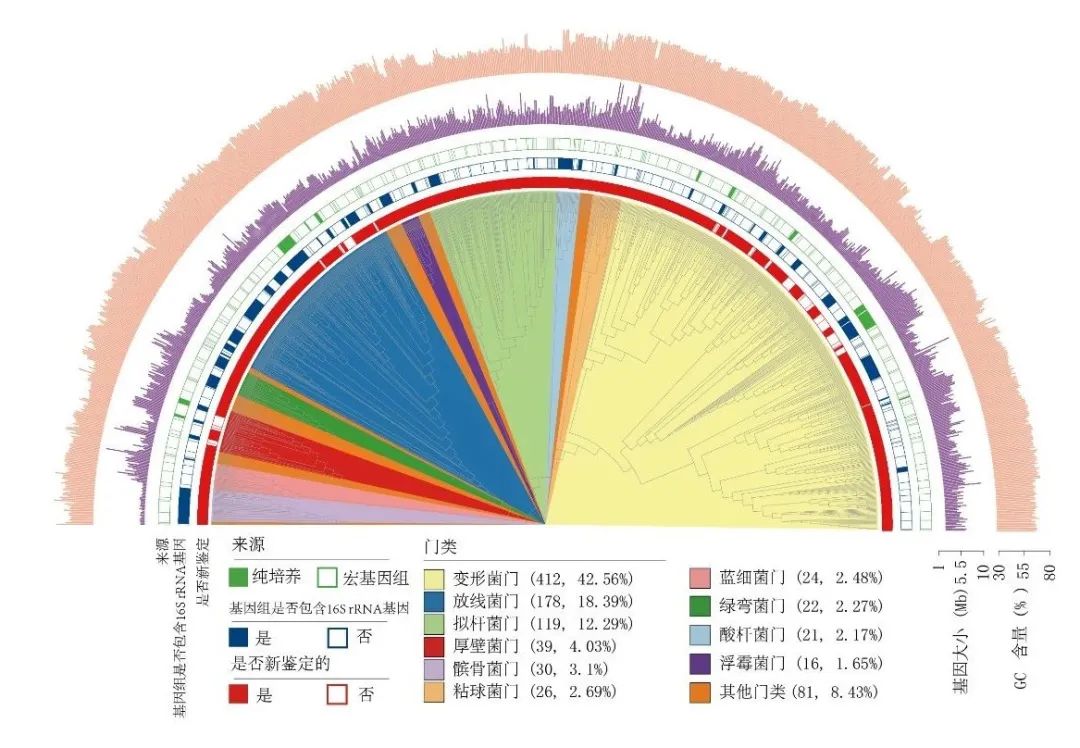
图8.获取的青藏高原冰川微生物968种细菌代表基因组系统发育树,图源:第二次科考队。
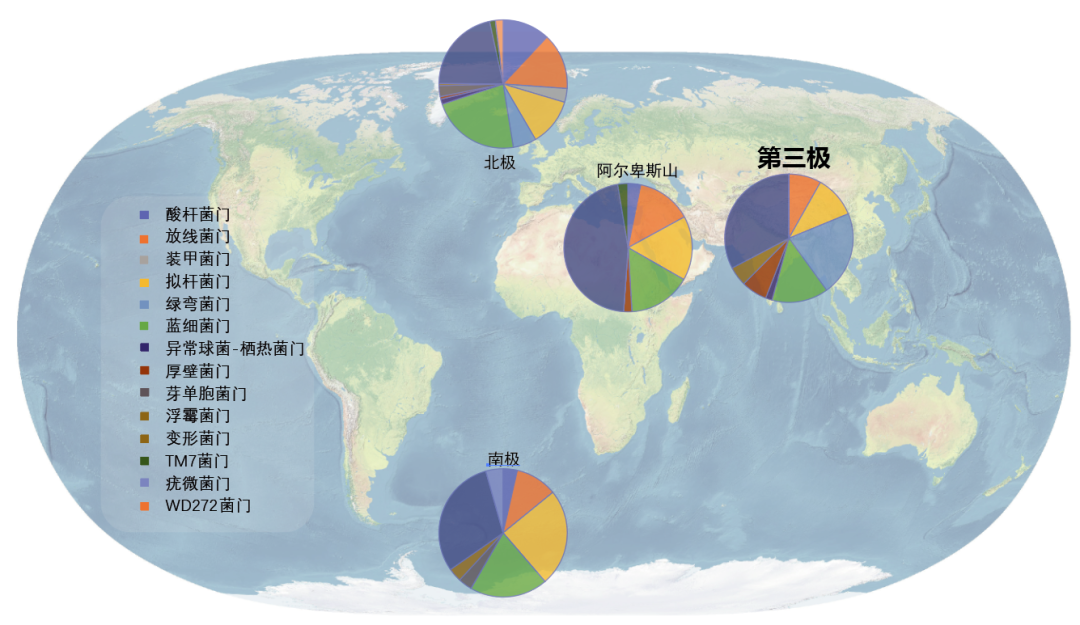
图9.青藏高原冰川微生物群落与其他地区冰川显著不同,图源:第二次科考队。
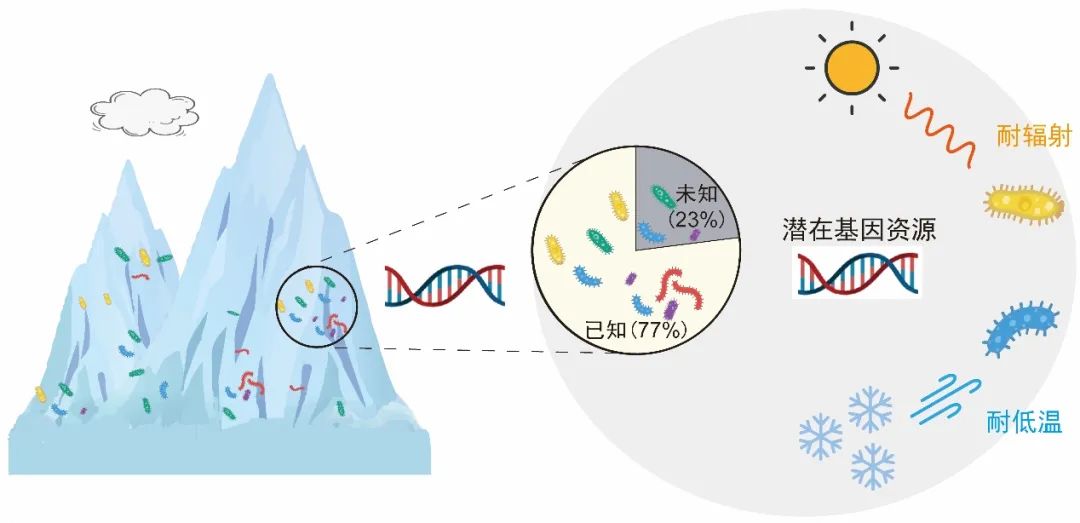
图10.青藏高原冰川蕴藏着独特的微生物,具耐低温和耐紫外辐射功能潜力,图源:第二次科考队。
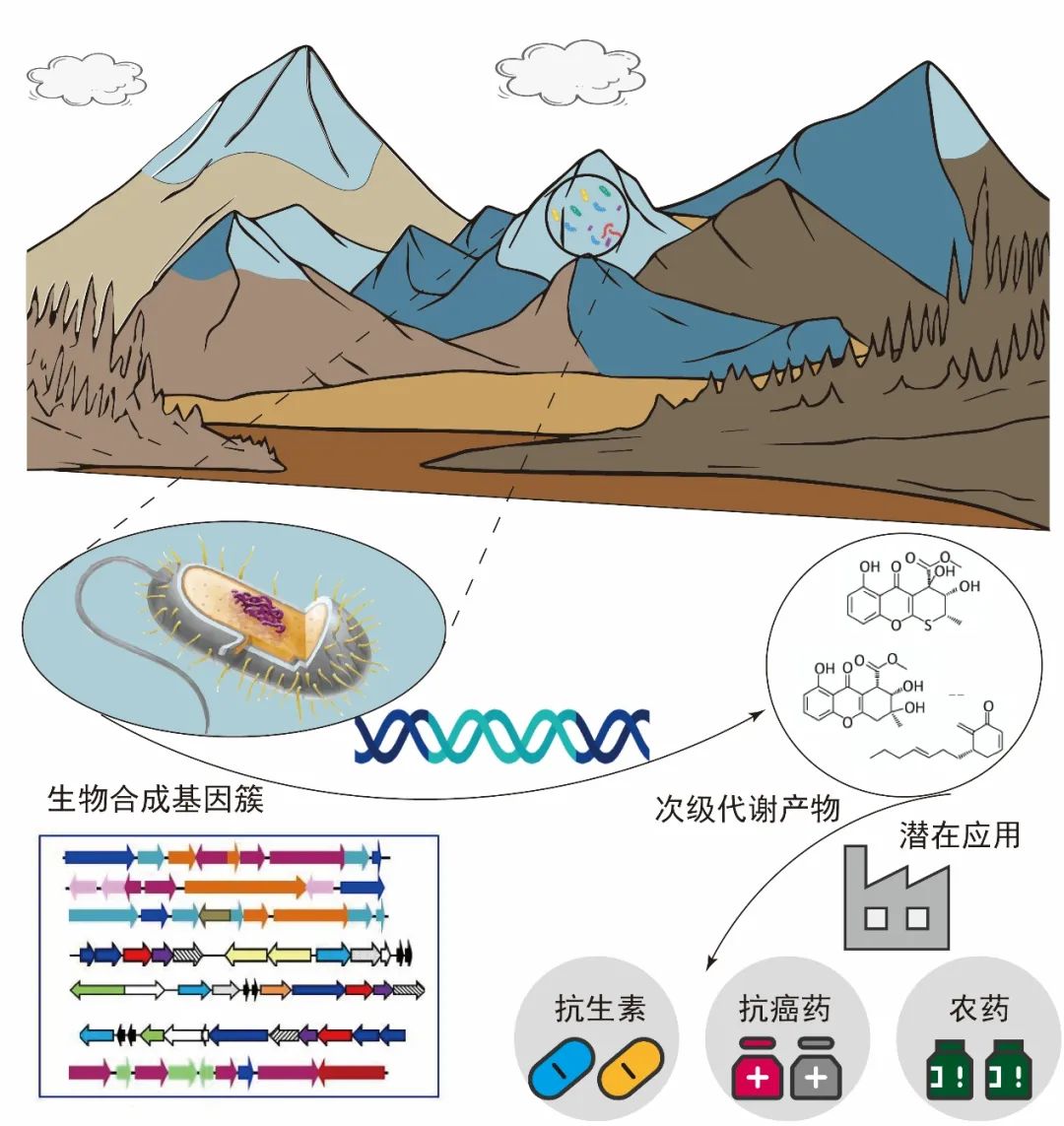
图11.青藏高原冰川微生物次级代谢产物的应用价值,图源:第二次科考队。
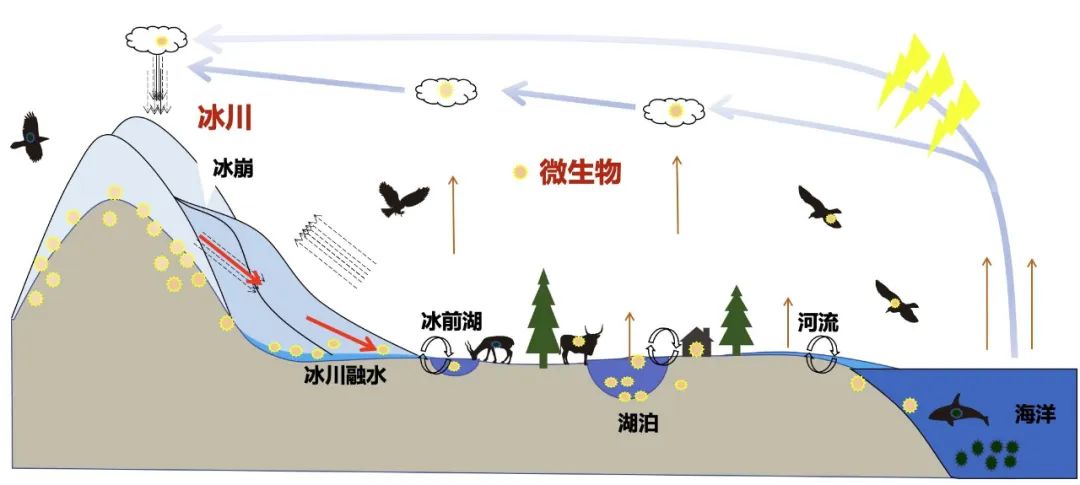
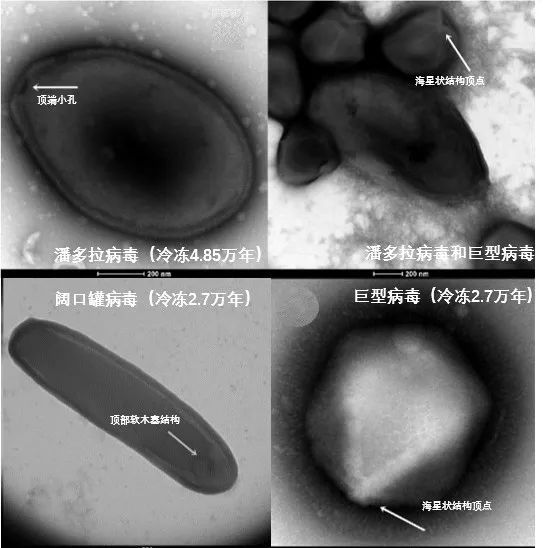
图13.科学家从西伯利亚永久冻土中复活的冷冻了数万年的病毒,图源:引自文献[17]。
2021年,美国俄亥俄州立大学伯德极地和气候研究中心报道,在青藏高原古里雅冰芯中发现了33个病毒物种,其中仅有12%为已知病毒,但它们主要为侵染细菌的噬菌体且数量极低。最近,兰州大学泛第三极环境中心团队研究了青藏高原26条冰川雪、冰和冰尘中的病毒,共发现了8894个病毒物种,种类数远低于土壤、湖泊和海洋等环境,其中西风区冰川中病毒数量大于季风区。尽管38%的病毒为新病毒种类,但以噬菌体为主,对人类无害。与人类相关病毒的相对丰度极低,仅百万分之一,且均为与人类长期共存、被人类熟知的病毒。因此,青藏高原冰川中的病毒对人类社会造成危害的风险极低。
在青藏高原冰川中,虽然与人类相关的病毒风险低,但是科考队员发现了微生物抗生素抗性基因的潜在危害。微生物利用其抗性基因,通过外排、解毒和靶点修饰等机制对抗抗生素,从而有利于微生物自身更好地适应环境,尽管微生物抗性基因的产生是一种自然和古老的过程。但在抗生素大量使用、环境浓度升高的背景下,这些抗性基因可通过水平基因转移在微生物间,及环境与微生物转移和传播,造成了耐药菌的滋生和扩散,甚至出现了能耐受几乎所有抗生素的“超级细菌”,导致重大的公共卫生危机。
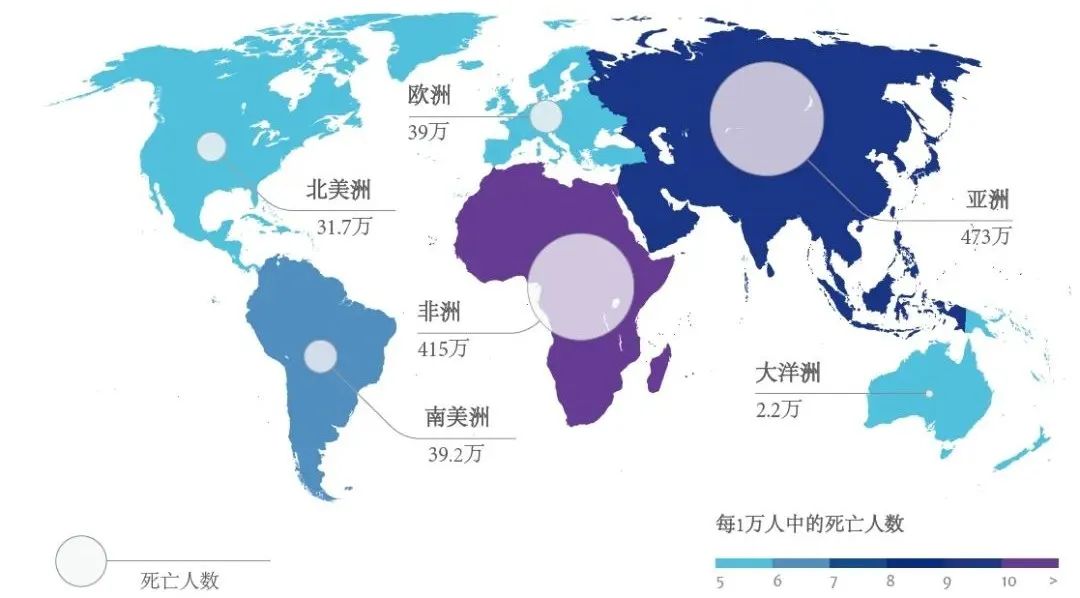
图14.全球预计到2050年每年因耐药性微生物感染致死人数,图源:The Review on Antimicrobial Resistance 2014

图15.2014年和2050年因耐药性微生物感染致死与其他主要死亡原因死亡人数对比,图源:The Review on Antimicrobial Resistance 2014。
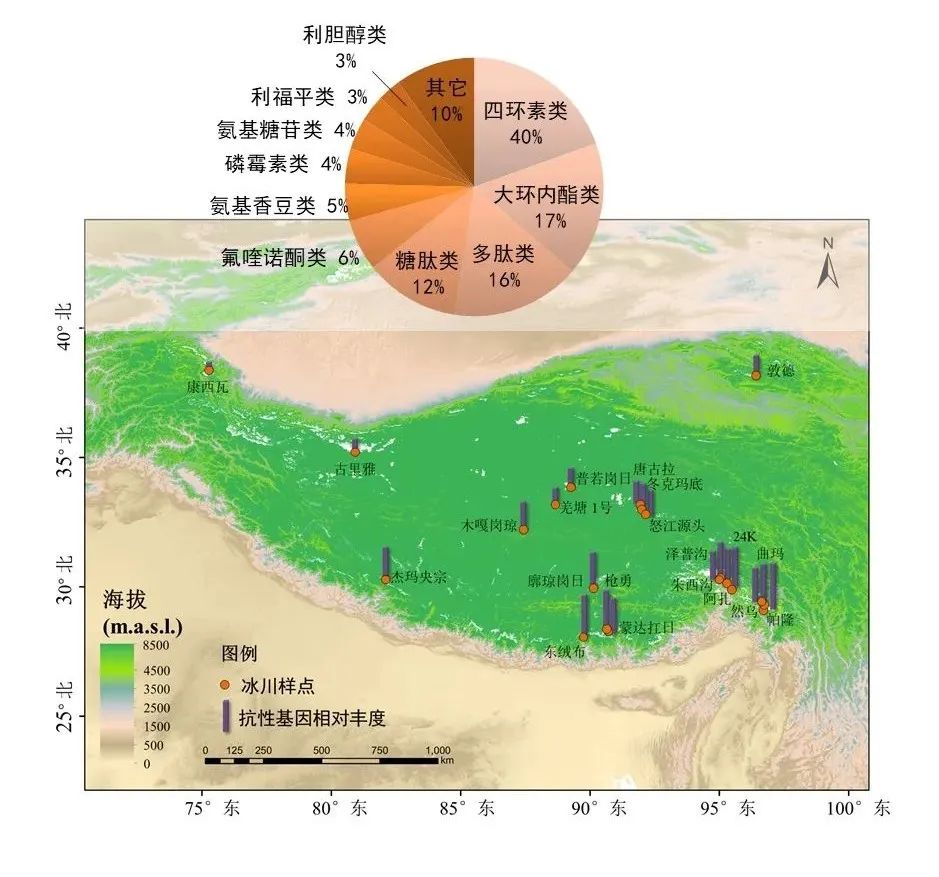
图16.青藏高原21条冰川抗生素抗性基因相对丰度以及主要抗性基因类型,图源:第二次科考队。
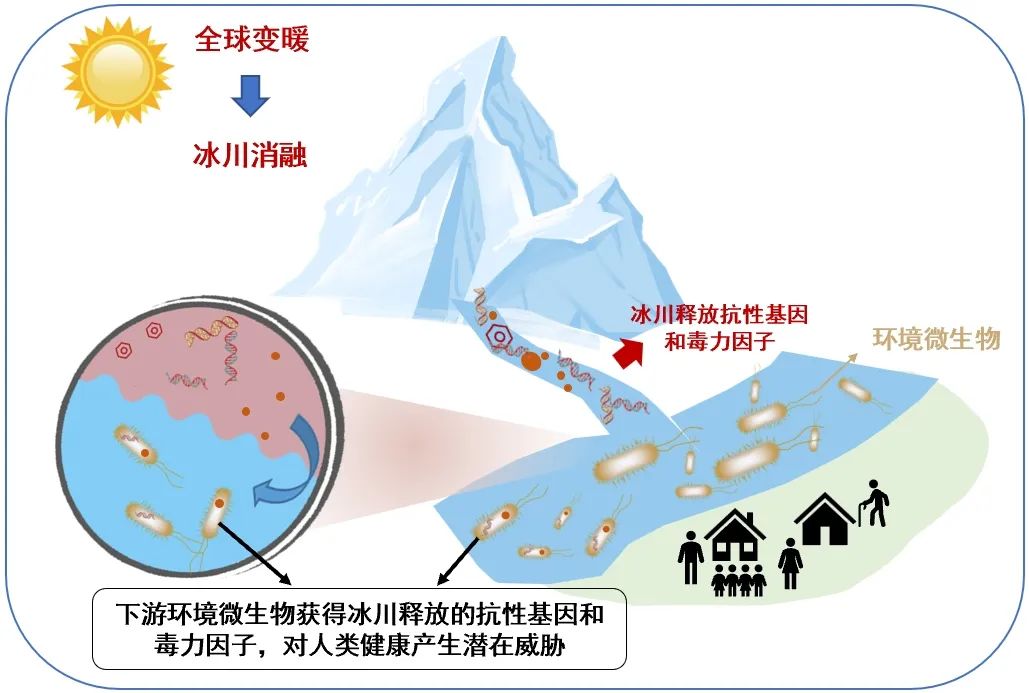
图17.抗性基因和毒力因子向冰川下游传播的示意图,图源:第二次科考队。
[1] Whittaker, R. H. 1969. New concepts of kingdoms or organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. Science, 163(3863):150-160.
[2] Woese, C. R. and Fox, G. E. 1977. Phylogenetic structure of the prokaryotic domain: the primary kingdoms. Proceedings of the National Academy of Sciences of the United States of America, 74(11):5088-5090.
[3] Shu, W.-S. and Huang, L.-N. 2022. Microbial diversity in extreme environments. Nature Reviews Microbiology, 20(4):219-235.
[4] Priscu, J. C., et al. 2007. Biological material in ice cores. Encyclopedia of Quaternary sciences, 2:1156-1166.
[5] Rogers, S. O., et al. 2004. Recycling of pathogenic microbes through survival in ice. Medical Hypotheses, 63(5):773-777.
[6] 刘时银 等. 2015. 基于第二次冰川编目的中国冰川现状. 地理学报, 70(01):3-16.
[7] 姚檀栋 等. 2019. “亚洲水塔”变化与影响. 中国科学院院刊, 34(11):1203-1209.
[8] Xiang, S. R., et al. 2005. Vertical quantitative and dominant population distribution of the bacteria isolated from the Muztagata ice core. Science in China Series D-Earth Sciences, 48(10):1728-1739.
[9] Yao, T., et al. 2012. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nature Climate Change, 2(9):663-667.
[10] Anesio, A. M., et al. 2009. High microbial activity on glaciers: importance to the global carbon cycle. Global Change Biology, 15(4):955-960.
[11] Anesio, A. M., et al. 2017. The microbiome of glaciers and ice sheets. NPJ Biofilms Microbiomes, 3:10.
[12] Anesio, A. M. and Laybourn-Parry, J. 2012. Glaciers and ice sheets as a biome. Trends in Ecology and Evolution, 27(4):219-225.
[13] 陈德亮 等. 2015. 青藏高原环境变化科学评估: 过去、现在与未来. 科学通报, 60(32):3025-3035.
[14] Liu, Y., et al. 2022. A genome and gene catalog of glacier microbiomes. Nature Biotechnology, 40(9):1330-1331.
[15] Revich, B. A. and Podolnaya, M. A. 2011. Thawing of permafrost may disturb historic cattle burial grounds in East Siberia. Global Health Action, 4.
[16]https://www.natureworldnews.com/articles/45267/20210222/prehistoric-animal-viruses-permafrost-caught-interest-russian- scientists.htm, Nature World News, feb 22, 2021.
[17]Alempic,J.M., et al. 2023. An update on eukaryotic viruses revived from ancient permafrost. bioRxiv, 2022.2011.2010.515937.
[18] Zhong, Z. P., et al. 2021. Glacier ice archives nearly 15,000-year-old microbes and phages. Microbiome, 9(1):160.
[19] 朱永官 等. 2015. 抗生素耐药性的来源与控制对策. 中国科学院院刊, 30(4):509-516.
[20] Wright, G. D. 2007. The antibiotic resistome: the nexus of chemical and genetic diversity. Nature Reviews Microbiology, 5(3):175-186.
[21] Liu, Y. Y., et al. 2016. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infectious Diseases, 16(2):161-168.
[22] Mao, G., et al. 2023. Monsoon affects the distribution of antibiotic resistome in Tibetan glaciers. Environmental Pollution, 317.